For the Scientists out there
A Redox Framing on Sirtuins, Yamanaka Factors, and Martin Feelisch

Before Watson and Crick, the heavy hitters in biology were focused on cellular metabolism. Seventy years later, we’ve seen that genetic and epigenetic factors are not the whole picture. The Human Genome Project was supposed to address the main factors of health and disease. It did not, ergo, the Human Exposome Project was launched to study the non-purely genetic factors of health and disease. There was considerable excitement this month about the recent heritability (genes plus other heritable factors) of longevity being calculated as up to 50% when you take out extrinsic causes of death.
I stumbled into longevity and the exposome research as an outsider. My primary expertise is in brain and metabolic imaging. I’ve developed hardware for magnetic resonance imaging and innovated clinical applications of imaging technology. I spent a period of two to three years diving deep into the medical literature based on an insight I had as a patient. First, I hypothesized that the glutathione redox status was a marker of neuroinflammation, which it appears to be. Then I realized that a lot of diseases that were coming up in the searches were those diseases typically associated with health disparity. So I looked into the hypothesis: diseases associated with health disparity present with low glutathione status (ratio of reduced to oxidized glutathione— GSH:GSSG) – which checked out for 100% of the diseases that I looked into. Then, I flipped the hypothesis: diseases with low glutathione status present as health disparities – which also checked out for 100% of the diseases that I looked at. Then I thought to myself, but this is looking like all of the diseases. So, I checked out each of Peter Attias’ horsemen: cardiovascular, metabolic, neurodegenerative, and cancer … yup, yup, yup, and yup. Glutathione redox status is also a measure of mitochondrial homeostasis, or resilience to stress. The mental health medical literature also suggested that glutathione status could explain why childhood traumas played a role in health and wellbeing later in life. And then I found Chris Palmers’ book, Brain Energy. In it he says, “all diseases are cross-correlated which statistically speaks to an upstream shared mechanism.” I think that’s the quote verbatim. His book speaks at length of his unified theory of mental illness. Then I went into the longevity literature and found that measures of glutathione were able to predict rate of disease accumulation over a 6-year period. Another paper showed that the centenarians they looked at had glutathione status more similar to that of middle-aged folks than seniors. Another study predicted telomere length from chronologic age and a measure of glutathione with r-values upwards of 0.8. I realized from what I read that biochemically (e.g., looking at p53) stress on healthy tissue upregulated resilience to stress, and that when stress fell on stressed tissue inflammation, disease, and aging followed. When reactive oxygen species build up, Kreb’s locks up at succinate and sends out hypoxia inducing factor one alpha initiating inflammation and switching into glycolysis – the less efficient though less endo-toxic form of ATP production. I read about redox proteomics--that redox status is also critical for the shape and function of our key proteins. And that stress signaling was a critical shared pathway in order to coordinate—as an organism—from cellular metabolism to behavior. That’s when I understood that the glutathione redox status was just a redox metabolic measure — something we should be tracking as a health marker, but also to prevent redox status as being a confounding variable.
At that point, I captured IP in the area of biomarkers of health. At this point, I am of the opinion that there is a Martin Feelisch sized hole in the existing models of Aging – and that adding the body’s redox architectures to the existing knowledge base shakes out a lot of the complicated disjointed aspects and better captures the complexity. Below are a couple references and images that speak to the role of redox in sirtuins, the Yamanaka factors, and Martin Feelisch’s work.
Sirtuins
Antioxid Redox Signal. 2018 Mar 10;28(8):643–661. doi: 10.1089/ars.2017.7290
The Role of Sirtuins in Antioxidant and Redox Signaling
Chandra K Singh 1, Gagan Chhabra 1, Mary Ann Ndiaye 1, Liz Mariely Garcia-Peterson 1, Nicholas J Mack 1, Nihal Ahmad 1,✉
https://pmc.ncbi.nlm.nihhttps://pmc.ncbi.nlm.nih.gov/articles/PMC5824489/.gov/articles/PMC5824489/
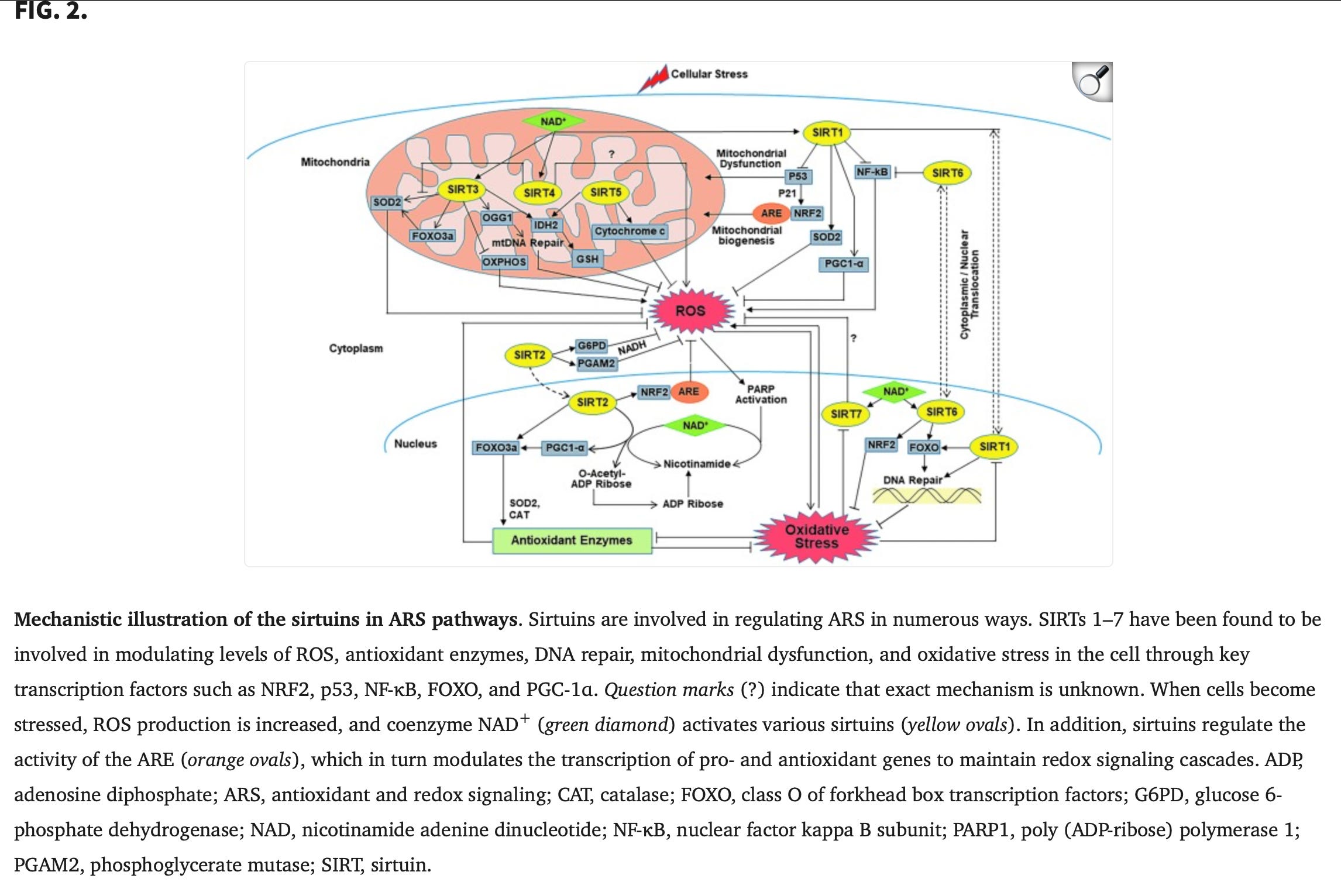
Yamanaka Factors:
Cysteine oxidation wildly alters OCT4 stability and function
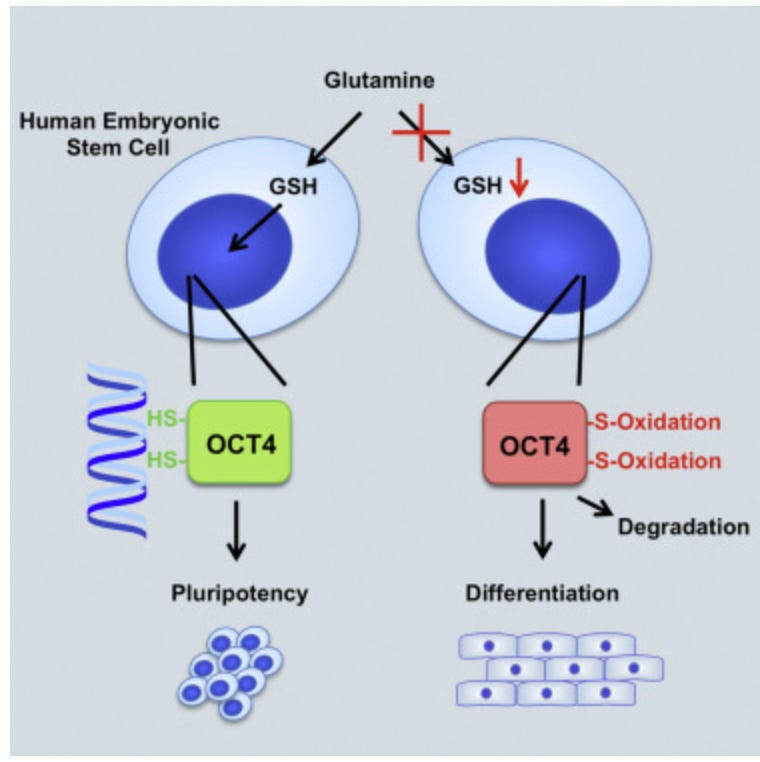
https://www.cell.com/cell-reports/fulltext/S2211-1247(16)30702-1
GSH = reduced glutathione
Likewise, Sox2 is very redox sensitive:
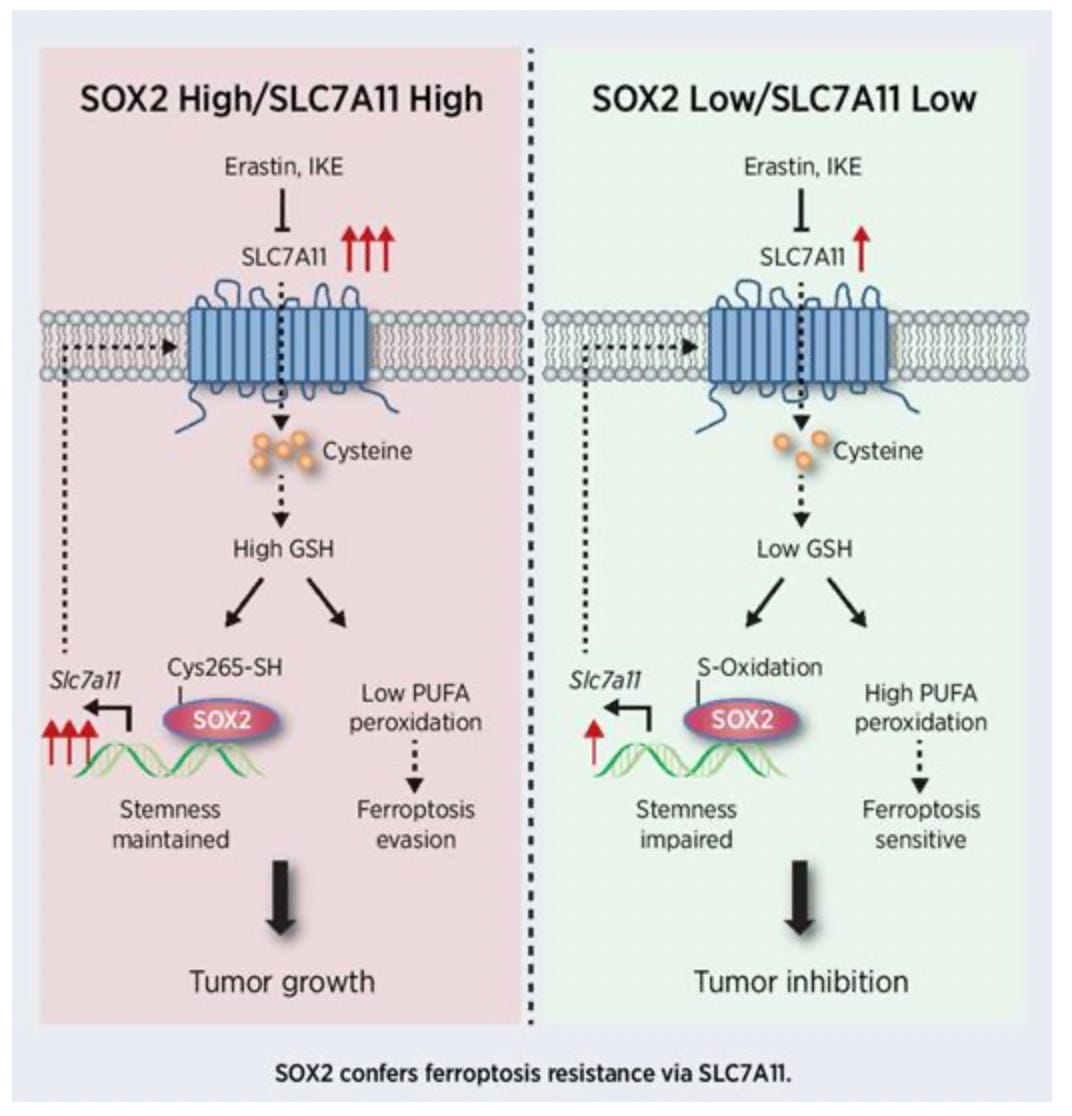
https://aacrjournals.org/cancerres/article-abstract/81/20/5217/670781/Stem-Cell-Factor-SOX2-Confers-Ferroptosis GSH = reduced glutathione
KLF4 is highly modified by the redox signaling/modification s-nitrosation, which is also reduced by glutathione upregulating compound NAC
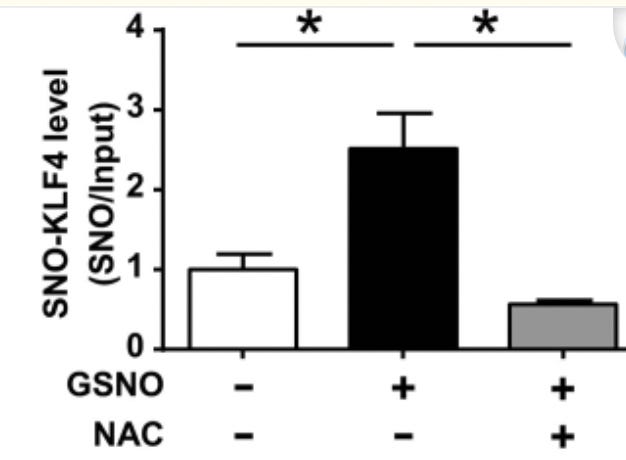
https://pmc.ncbi.nlm.nih.gov/articles/PMC6348764/
NAC increases GSH
In hypoxia-induced rat model of pulmonary arterial hypertension (PAH), S-nitrosated KLF4 (SNO-KLF4) was significantly increased in lung tissues, along with decreased nuclear localization of KLF4.
Here, we demonstrated that S-nitrosation (GSNO) treatment is a novel mechanism to post-translationally modify KLF4 activity. Such a modification can impair the vasoprotective function of KLF4 and may be implicated in the pathogenesis of PAH.
Redox regulation of the embryonic stem cell transcription factor oct-4 by thioredoxin
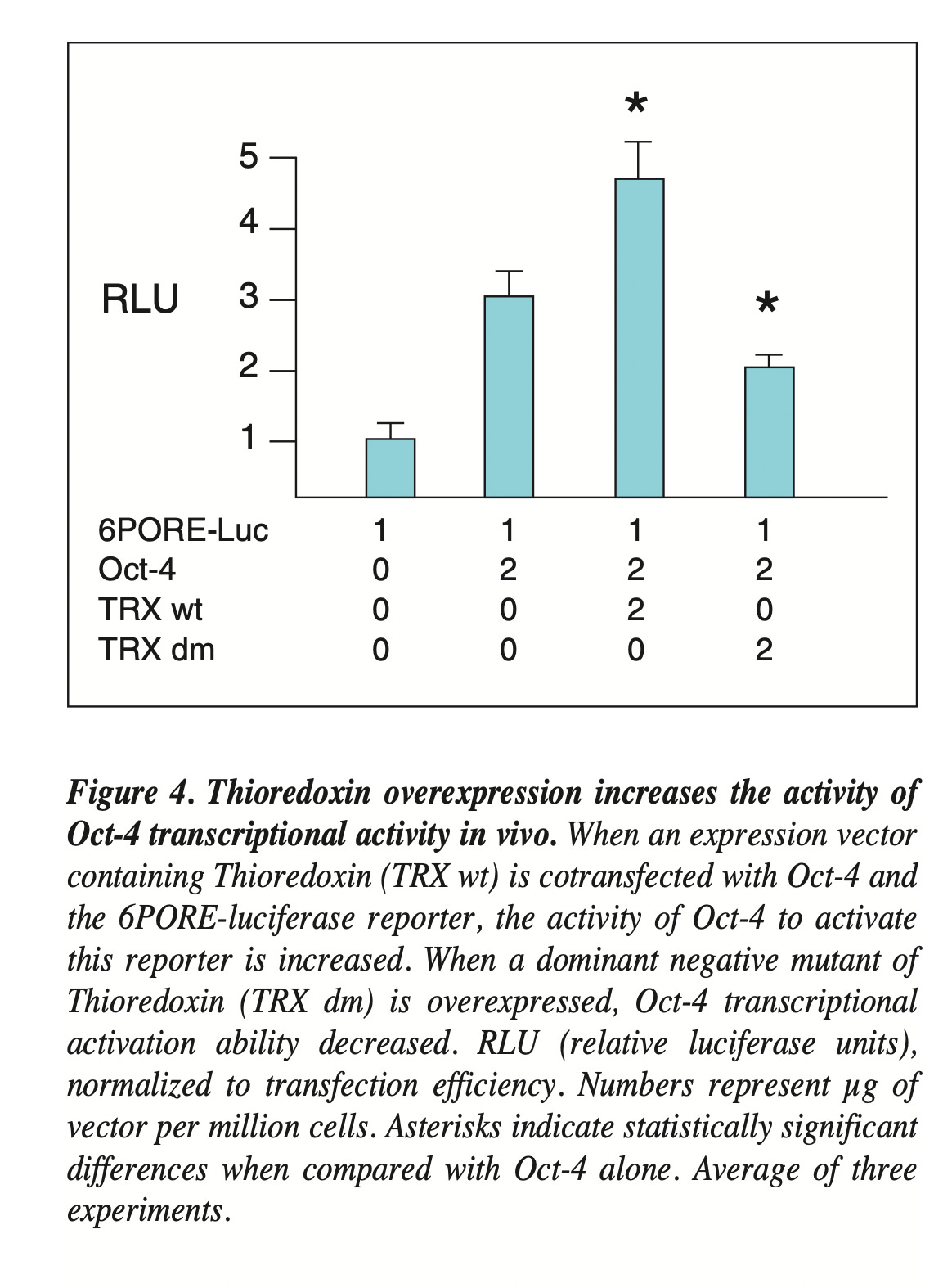
https://pubmed.ncbi.nlm.nih.gov/15153603/
Oct4 redox sensitivity potentiates reprogramming and differentiation
Genes Dev
. 2024 Apr 1;38(7-8):308–321. doi: 10.1101/gad.351411.123
“Here, we pinpoint Oct4 redox sensitivity as an important determinant of both reprogramming and differentiation”. – and it comes down to a single cysteine (w/ neighboring arginine.
Martin Feelisch’s Work
. 2022 Mar 21;21:623–646. doi: 10.17179/excli2022-4793
Systems redox biology in health and disease
Martin Feelisch 1,*, Miriam M Cortese-Krott 2, Jérôme Santolini 3, Stephen A Wootton 4, Alan A Jackson 4
Abstract
Living organisms need to be able to cope with environmental challenges and other stressors and mount adequate responses that are as varied as the spectrum of those challenges. Understanding how the multi-layered biological stress responses become integrated across and between different levels of organization within an organism can provide a different perspective on the nature and inter-relationship of complex systems in health and disease. We here compare two concepts which have been very influential in stress research: Selye’s ‘General Adaptation Syndrome‘ and Sies’s ‘Oxidative Stress‘ paradigm. We show that both can be embraced within a more general framework of ‘change and response’. The ‘Reactive Species Interactome‘ allows each of these to be considered as distinct but complementary aspects of the same system, representative of roles at different levels of organization within a functional hierarchy. The versatile chemistry of sulfur - exemplified by hydrogen sulfide, glutathione and proteinous cysteine thiols - enriched by its interactions with reactive oxygen, nitrogen and sulfur species, would seem to sit at the heart of the ‘Redox Code’ and underpin the ability of complex organisms to cope with stress.
1. Oxidative Stress and Redox-Modulating Therapeutics in Inflammatory Bowel Disease
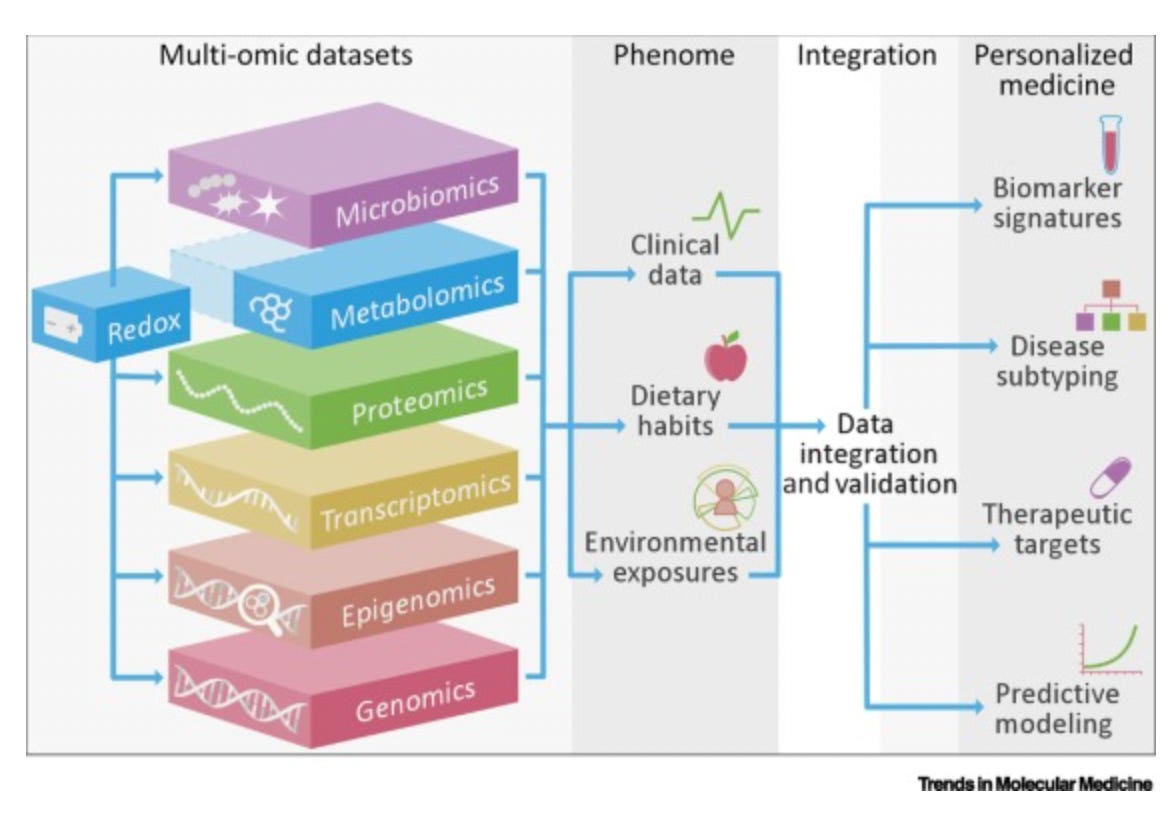
Bourgonje, Arno R. et al. Trends in Molecular Medicine, Volume 26, Issue 11, 1034 - 1046
The Reactive Species Interactome: Evolutionary Emergence, Biological Significance, and Opportunities for Redox Metabolomics and Personalized Medicine
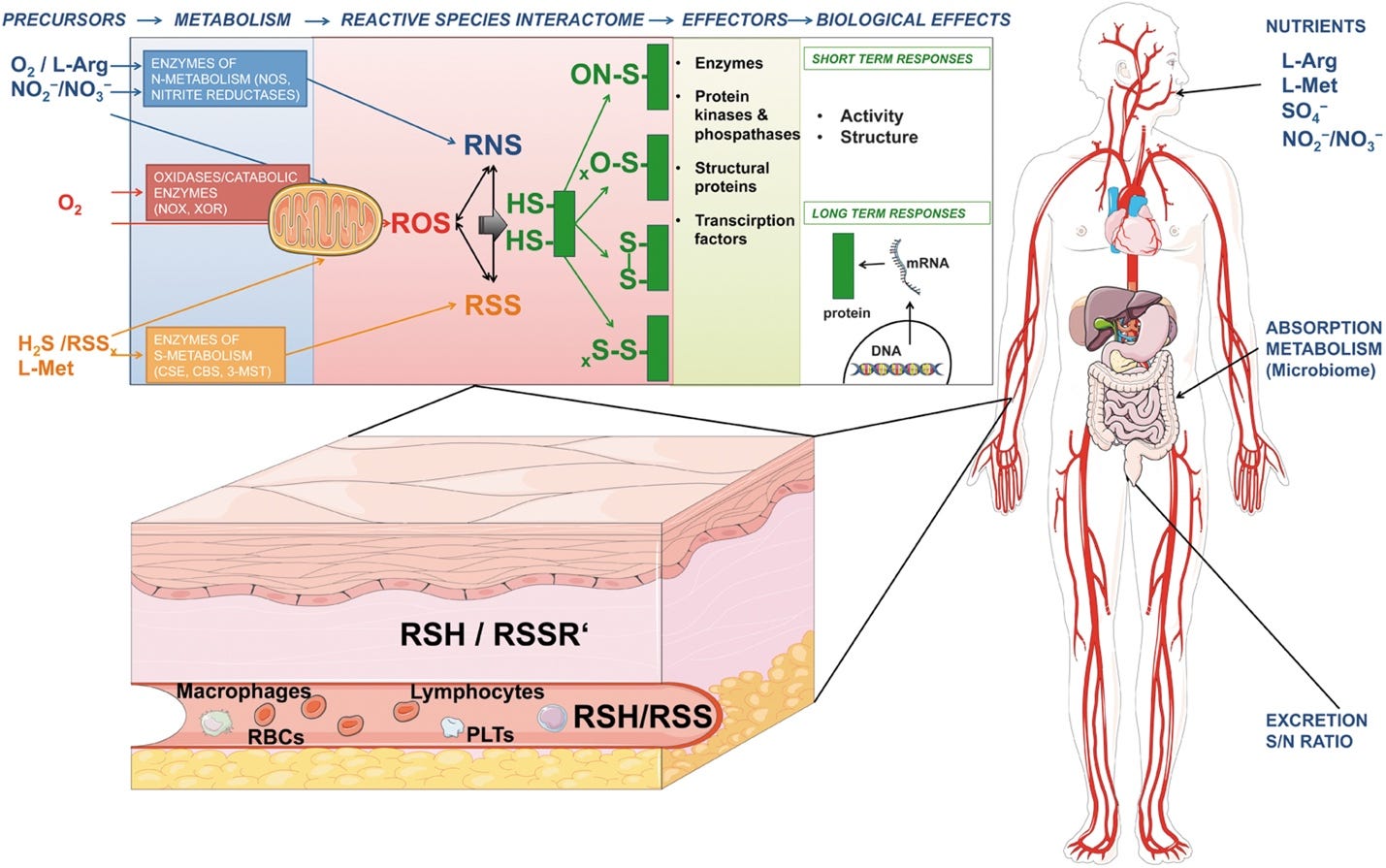
https://journals.sagepub.com/doi/full/10.1089/ars.2017.7083
FIG. 1. Intracellular, extracellular, and interorgan/systemic role of the RSI. Precursors of the RSI are organic and inorganic substrates and cofactors, including amino acids (e.g., arginine, methionine), vitamins (B6, B12, C), and xanthine, as well as oxygen, nitrite, polysulfides, thiosulfate, and sulfate, which are transformed by mitochondrial or cytoplasmic enzymes into ROS, RNS, and RSS. The chemical interactions among ROS, RNS, and RSS lead to formation of a number of products with different reactivities, stabilities, half-lives, and therefore different lifetimes defined by their physicochemical properties, covering a wide range of maximal travel distances. A common target of the RSI are cysteine thiols in proteins, acting as redox switches, able to fine-tune activity of signaling molecules, and leading to short-term responses (e.g., protein kinases and phosphatases inducing changes in signaling and glucose metabolism) or long-term adaptation (by modifying redox switches responsible for gene expression regulation, such as the HIF, NFkB, and Keap1/Nrf2 pathways). The RSI serves also as a local and systemic heterocellular communication system mediated by actions of longer-lasting products of the RSI (e.g., nitrite, polysulfides) and circulating thiols. The nutritional and physiological status of the organism affects the RSI by reciprocally regulating precursor availability, metabolism, signaling, and mitochondrial function. RNS, reactive nitrogen species; ROS, reactive oxygen species; RSS, reactive sulfur species.